
阴还是阳: 窦房结起搏机制的中医哲学


放一篇文章在这,如有侵权,请与本人联系,谢谢。
引用:王泰一, 姚茜, 解微微, 等. 阴还是阳: 窦房结起搏机制的中医哲学[J]. 中南药学, 2017, 15(3): 257-263.
阴还是阳:窦房结起搏机制的中医哲学
王泰一1,2,姚茜1,2,解微微1,2,崔英1,2,朱彦1,2*(1. 天津中医药大学,天津市现代中药国家重点实验室,天津 300193;2. 天津国际生物医药联合研究院,中药新药研发中心,天津 300457)
摘要:中医药治疗心律失常已有数千年的历史,但抗心律失常中药药理的研究过程中往往仅能参考西药药理学机制开展研究,而对于中药所特有的“理性”的多靶标调控机制则缺乏相应的理论探索。本文通过回溯窦房结起搏机制的研究历史,结合中医阴阳理论解析和重述起搏机制中离子通道及信号转导机制,探讨窦房结起搏机制的阴阳变化,以期从中医角度定义窦房结起搏机制的物质基础,希望能为中医药治疗窦性心律失常提供更多地思路。
关键词:窦房结;起搏机制;中医哲学;动作电位;离子通道
阴阳是中国古代哲学的一对范畴。阴阳的最初涵义很朴素,表示阳光的向背,向日为阳,背日为阴,后来引申为气候的寒暖,方位的上下、左右、内外,运动状态的躁动和宁静等。中医理论体系中引入了一分为二的阴阳辩证观,用来观察天、地、人,尤其用来观察人的形体结构、生理功能、病理变化,乃至辨证治疗,用这种辩证思维模式,可以居安思危、治病防病、治已防传、祛邪护正,这无疑更具科学性[1]。现代生物化学、细胞生物学发源以来,各种概念理论与中医传统理论逐渐发生碰撞和交汇,传统中医理论也在吸纳现代科学的同时得到进一步的延伸和拓展。
本文试图从阴阳学说角度结合现代生物学理论探讨窦房结起搏机制、病态窦房结综合征的病理机制及治疗。窦房结位于上腔静脉与右心耳之间的心外膜深面,呈椭圆形,是高等脊椎动物心脏的主要起搏细胞,它的起搏节律决定整个心脏的节律[2]。中医认为,心为君主之官,主一身之血脉,心阳鼓动心脏搏动,推动全身的血脉循行;而肾阳为诸阳之本,又从更加根源的角度影响着心阳,因此,心肾阳气的盛衰,直接影响心率的快慢、血脉的盈亏和脉象的虚实[3]。而随着年龄的增长,心肾阳气逐步走向衰竭,于心脏起搏组织中的表现即为窦房结中离子通道的表达量下降,进而造成了起搏功能的降低[4]。
一、起搏机制研究进展
由于窦房结体积较小,不同部位细胞异质性较高,起搏机制研究方面一直面临着较大的困难,研究进展缓慢而且争论不断,不同时期有不同的争论焦点[5]。
最初,人们往往希望找到一种关键性的起搏电流,或离子通道,来阐明起搏机制。在funny电流发现以前,比较公认的起搏机制是心脏起搏细胞的去极化过程由一种衰减的外向钾电流引发,1968年, Noble和Tsien在Purkinje纤维中进一步记录到了这种钾通道IK2[6]。由于IK2的反转电位在钾离子的平衡电位附近,因而被描述为一种只对钾离子通透的离子通道。此后又有研究发现,IK2还参与了交感神经激动所引发的心率上升过程,于是这种“衰减的外向钾电流”理论得到了前所未有的认可,尽管这一理论并不能解释全部的现象,但由于来自于确凿的实验证据,故而统治了起搏研究领域10余年之久[7-9]。
直到1979年,DiFrancesco 等在窦房结中发现了一种 “有趣”的内向电流,这种电流由超极化激活,也是窦房结起搏的机制之一,二者的理论冲突从此产生:窦房结和Purkinje纤维同属于心脏传导系统,如此高度关联的两个组织竟然同时存在两种“对立”的起搏机制[10]。但这种对立并未存在太长的时间,1981年,DiFrancesco 等再次证明以往发现的IK2 实际是实验中If与IK1两种通道共同形成的假象,从而彻底否定了“衰减的外向钾电流”理论[10-12]。此后,If电流的研究进入了一个新的阶段,对于起搏电流的探索也集中到了对于内向电流的研究上[13-17]。基因表达研究发现,funny电流由超极化激活环化核苷酸门控通道(Hyperpolarization-activated Cyclic Nucleotide-gated,HCN)所产生[18]。HCN的4种亚型中,心脏起搏细胞中HCN4通道对funny电流贡献最大,而HCN1和HCN2表达量较低[19,20]。为了验证funny电流能够主导自律性的起搏功能,Qu等[21]在犬的左心房转染HCN2基因,使心房局部高表达funny current,结果显示犬的心房可以形成具有自律性的起搏位点。然而,Noma等研究[22]认为,DiFrancesco发现的If电流在动力学上表现过于缓慢而且通道激活时的电压太低(<-70 mV),生理条件下的电流也仅为0.5~2 pA,尚不足以主导起搏的发生。1995年,Noma团队报道了一种新发现的电流Ist,这是一种L型钙通道的新亚型,能够被有机的和无机的Ca2+通道阻断剂所阻断,而对于钠通道阻断剂河豚毒素(tetrodotoxin,TTX)不敏感[22]。在20世纪80年代至90年代初,尽管对于起搏电流的争论从未停止过,但研究者一致认同起搏电流应当是一种“内向”的离子流,才能导致细胞去极化的发生。
1996年,上述细胞膜内向电流引发去极化发生这一论调尚未尘埃落定之际,Rigg等的研究又将起搏机制引向了另一个方向——内钙释放[23],Ryanodine阻断肌浆网的钙释放之后,心率随之下降。随后,Huser等[24]在猫心房起搏细胞中进一步证实了细胞内钙释放对于自律性产生的必要性。然而,仅仅内钙释放并没有产生跨膜的离子流动,也就是说缺乏一个显著的内向电流引发去极化。Ju等[25]的工作使这一理论得以完善:肌浆网内钙释放导致的细胞质Ca2+浓度快速升高,进而激活钠钙交换体(Na+-Ca2+ exchanger,NCX)将Ca2+排向细胞外,由于钠钙交换体的Na+-Ca2+交换比为3:1(Na+内流的电流为Ca2+外流的3倍),因而钠钙交换体产生的净电流为内向电流,具备了引起细胞去极化的条件。
上述两类起搏机制理论表明,起搏细胞发生去极化至少存在这样两种机制:(1)电压依赖的跨膜内向离子流(如If,Ist等等);(2)肌浆网Ca2+释放激活钠钙交换体,引发起搏细胞去极化[2]。近年来,Lakatta 实验室综合10 多年的研究成果,提出膜时钟和钙时钟相耦联的“双时钟”学说[26],即细胞膜上各种离子流周期性的活动(膜时钟)与细胞内肌浆网Ca2+周期性的释放(钙时钟)相偶联,虽然钙时钟具有自发的节律变化,但膜时钟在每个动作电位周期中反复校准钙时钟的节律,从而达到高度同步的状态[27],如图1所示。
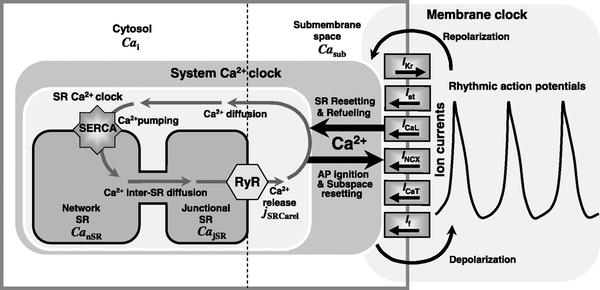

图1 膜时钟和钙时钟相耦联的“双时钟”起搏机制[26]。细胞膜上的L型钙通道诱发肌浆网的钙释放,而大量的Ca2+又激活细胞膜上的NCX,从而使细胞膜上的Na+、K+、Ca2+等离子通道形成了周期性的动作电位变化与肌浆网内钙释放的节律得到同步,形成偶联的“双时钟”。
二、起搏机制的阴阳理论
窦房结的起搏机制调控中涉及到一系列的离子通道、受体和第二信使,其中离子通道是窦房结电活动的直接承担者,不仅起到形成动作电位、传播起搏电信号的作用,也是窦房结规律性电位变化节律的调控者,而窦房结细胞的膜受体及其下游的信号转到分子则起到间接调控离子通道活性,控制起搏频率高低的作用。
1. 离子通道的阴阳属性
从“双时钟”理论中我们不难看出,在细胞膜与肌浆网同时存在着两种“循环”——细胞膜电位周而复始的升降变化的循环与细胞内肌浆网的Ca2+释放-回收的循环。就“形式”而论,膜电位的升高与降低可以视为一次阴阳变化,而Ca2+的释放与回收在细胞质内也同样表现为Ca2+浓度的升高与降低,这一出一入同样可以视为阴阳的变化。从物质的运动属性出发,趋向于“动”者为阳,趋向于“静”者为阴。对于窦房结起搏细胞来说,从动作电位(AP)角度看,去极化为阳,复极化为阴;从膜电位角度看,电位正向变化者为阳,负向变化者为阴;从离子通道电流方向角度看,内向通道为阳,外向通道为阴,起搏细胞动作电位及主要电流的阴阳属性如图2所示。
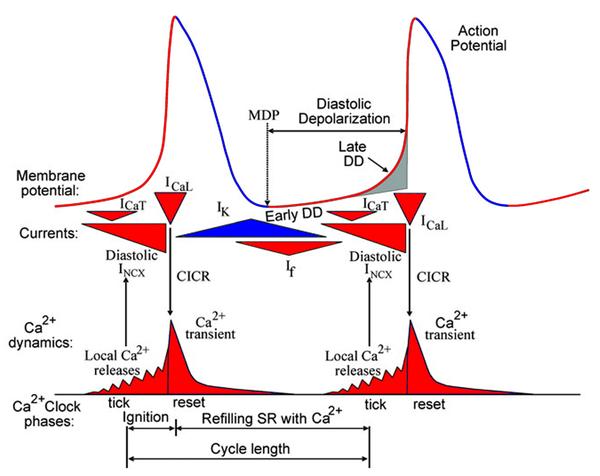

图2 起搏细胞动作电位及主要电流的阴阳属性。上部为起搏细胞动作电位曲线,中部为膜时钟内主要通道电流出现的时间及基本属性,下部为肌浆网钙时钟节律。图中红色表示属性为阳,蓝色表示属性为阴。本图由Lakatta等[27]发布,经作者修改。
然而,从动作电位时相考察仅能反应起搏细胞总体性的阴阳状态,并不足以定义每种离子通道的属性。作为一大类细胞内的功能蛋白,离子通道可以至少在两方面定义其属性——“体”和“用”,顾名思义,“体”即为离子通道的“器质”属性也就是说离子流的方向,依据前文的阴阳定义,电流方向指向细胞质(向细胞质积聚电荷)则为阳,指向细胞外机制或肌浆网则为阴。而“用”即为离子通道的“功能”属性,也就是从心脏起搏频率的角度考察其属性,增大离子通道的电流使起搏细胞APD缩短或者频率加快则为阳,反之则为阴。为了探讨窦房结pacemaker细胞起搏机制中的各种功能蛋白以及中药调节窦性心律的可能机制,我们参照运动属性的阴阳划分,重新定义了各种离子通道的属性,如表1所示。
表1 起搏机制中各离子通道的阴阳属性
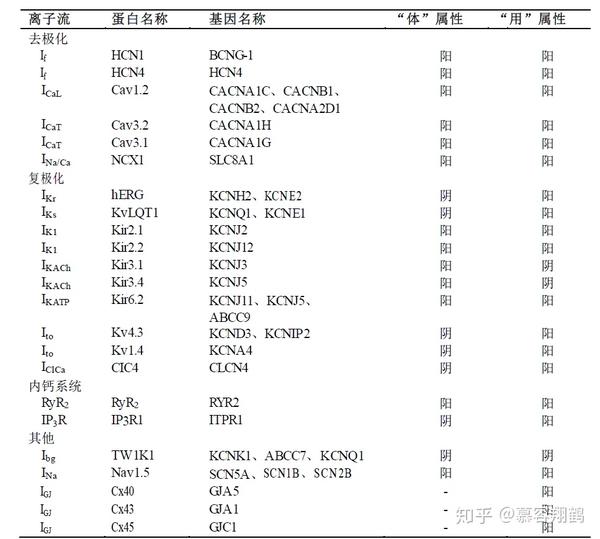

在起搏细胞去极化过程中,总电流方向为内向,首先funny电流(If)在4相早期使起搏细胞膜电位缓慢升高,而后T型钙通道(ICaT)和L型钙通道(ICaL)依次开放,伴随着肌浆网RyR2的开放产生0相去极化[28-34]。随着细胞内Ca2+浓度的上升,钠钙交换体(INa/Ca)被激活,它在向细胞外泵出1个Ca2+的同时向细胞内泵入3个Na+,因而其总电流方向为内向,起到促进细胞去极化的作用[35]。另外,研究表明钠钙交换体抑制剂能够显著延长APD,这也说明了钠钙交换体功能属性为阳,对它的阻断具有一定程度降低心率的作用[36]。上述各通道电流方向指向细胞质,通道的开放促进了起搏细胞去极化,因而器质属性和功能属性皆为阳,故为“体阴而用阳”。
在复极化过程中离子通道种类繁多,总电流方向为外向,但涉及到多种内向、外向电流。快速激活的延迟整流钾通道(IKr)、 缓慢激活的延迟整流钾通道(IKs)为复极化过程中的主要电流,两种电流均为外向整流通道[37,38]。起搏细胞中的瞬时外向钾电流(Ito)与心室肌中类似,激活与失活电压分别为+11 mV与-49 mV[39]。Ito抑制剂4-AP能够显著减少Ito电流,延长APD [40]。ICl,Ca通道为Ca2+激活的Cl-通道,通道的激活与肌浆网的Ca2+释放同步,通道的I-V曲线呈钟形,在+40 mV处达到峰值[41]。由于ICl,Ca通道为阴离子通道,故而其电流方向与离子流方向相反,又由于通道电流主要存在于快速复极的1相,其通道的开放主要起到缩短动作电位时程的作用。故而上述通道为“体阴而用阳”。在心衰的心脏中IK1的表达量被下调,而过表达IK1则会显著缩短APD加速3相复极[42]。KATP为ATP门控的钾通道,在缺氧条件下ATP浓度下降时通道打开,缩短APD加快心率[43]。上述2类通道电流方向向内,为“体阳而用阳”。在起搏所涉及的离子流中,IK, Ach通道为胆碱能受体偶联的钾通道,胆碱能受体的激活能够使通道打开,进而起到降低心率的作用[44],因而其属性为“体阳而用阴”。
在肌浆网内钙系统中,RyR2主要负责将肌浆网中的Ca2+释放到细胞质中,而IP3R则起到相反的作用[45]。由于钙瞬变与去极化高度同步,细胞膜去极化时RyR2快速释放Ca2+,而复极化时RyR2逐渐关闭,IP3R介导的Ca2+回流占据主导。因而,RyR2的作用相当于去极化膜离子通道的内向离子通道,故而定义为“体阳而用阳”;IP3R则相当于复极化时外向离子通道,因而为“体阴而用阳”。
此外, Ibg、INa、IGJ虽然并非起搏机制中的主要通道电流,但却与起搏电信号的维持和传播等功能密切相关。背景钾电流(Ibg)主要参与维持静息电位,通道的开放使细胞膜对K+维持一定的通透性,若减小该通道电流(如低钾条件下)细胞静息电位将正向移动,加速去极化。因此Ibg通道属性为“体阴而用阴”。与心房心室肌细胞不同,INa通道在窦房结中并不是主要的去极化电流,突变会导致病态窦房结综合征和传导障碍[46],故而我们依然将其属性归为“体阳而用阳”。在窦房结中,IGJ电流主要由Cx40、Cx43和Cx45承担,起到传导起搏电信号的作用。其中Cx40与 Cx45的表达量较为稳定,不随年龄的增长而变化,Cx43已证实与老年窦房结功能减退有关[47]。IGJ电流属于细胞间通讯连接,其电流方向具有双向性,因而其自身不存在阴阳属性,但它的表达量与起搏信号在心脏传导组织的扩散有关,因而其功能属性为阳。
2. 受体及信号转导分子的阴阳属性
窦房结起搏频率主要受自主神经系统的调控。β肾上腺素能受体是一种G蛋白偶联受体,其下游与Gs蛋白偶联,β1亚型在心脏中高表达。β1受体的激活能够增加心输出量,主要表现在增加心率、提高心肌收缩力、增强心脏自律性、提高传导速度。β1受体通过激活腺苷酸环化酶(AC)使细胞内cAMP浓度上升,进而增大funny电流幅度统计激活PKA,PKA则能够激活膜时钟的ICaL、IKr、IKs与钙时钟的RyR2和SERCA,加快动作电位的去极化和复极化,缩短动作电位时程。而M2胆碱能受体则起到相反的作用,通过抑制AC的活性降低上述通路下游分子的活性,使动作电位时程延长,心率减慢。而在GPCR-cAMP-PKA通路中也存在着负反馈机制,如PKA可以激活磷酸二酯酶PDE与蛋白磷酸酶PP,PDE能够降低cAMP浓度而PP则与PKA的底物分子存在广泛的去磷酸化作用。故而,β1受体、AC、cAMP、PKA的属性为阳,M2受体、PDE、PP的属性为阴。
此外,Ca2+作为一种第二信使也可以激活下游多种信号分子,例如钙调蛋白CaM能够激活AC进而引发PKA通路的反应,而CaMKII则对ICaL、ICaT、RyR2和SERCA具有激活作用,Ca2+自身也能够激活IClCa等钙依赖的离子通道。因而CaM和CaMKII的属性均为阳。
综合上述各离子通道及信号分子的属性,我们为起搏机制的通路每一环节定义了阴阳属性,如图3所示。
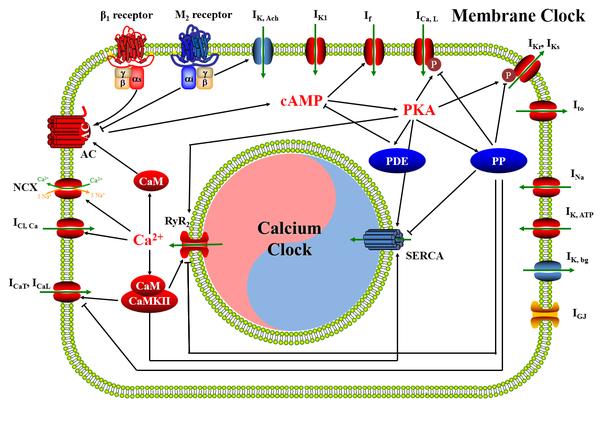

图3 窦房结起搏的阴阳变化机制。图中红色表示靶标的功能属性为阳,蓝色表示功能属性为阴,连线中箭头表示促进或激活关系,平头表示抑制作用。
三、窦性心律失常的中医药治疗
1. 参仙生脉口服液
参仙升脉口服液是由红参、淫羊藿、补骨脂、枸杞子、麻黄、细辛、丹参、水蛭共8味中药组成的复方制剂。侯平等[48]研究发现参仙生脉能使大鼠血清内的cAMP 显著升高是其提高窦性心律的药理机制之一。同样表现出多靶点效应。Jing等[49]发现,参仙升脉中麻黄的主要成分麻黄碱都能激活体外表达的KCNQ1/KCNE1通道,从而激活IKs电流,使窦房结自律性得到提升。
2. 麻黄附子细辛汤
麻黄附子细辛汤由麻黄、附子、细辛3味中药组成。张水馨等[50]研究发现,麻黄附子细辛汤治疗后心脏组织中CaM、CamK II蛋白表达量呈显著性上升。附子附子主要成分乌头碱具有显著的电生理效应,不仅能够增大钠通道(INa)的电流,对于IK1和ICaL也有显著地增强作用[51]。
3. 稳心颗粒
稳心颗粒由党参、黄精、三七、琥珀、甘松等5味道地中药材组成 [52]。稳心颗粒临床除了应用于早搏与室颤的治疗,也常用于窦性心动过速的治疗[53-58]。Chen等[59]研究发现稳心颗粒能够加速显著ICaL的失活并且延长通道恢复到静息状态的时间,5 g·L-1浓度的稳心颗粒即可显著抑制ICaL。Antzelevitch实验室[60] 与Wang等[61]均发现稳心颗粒能够对Nav1.5钠通道形成显著的使用依赖性抑制。此外,Xing等[62]研究发现稳心颗粒能够显著下调CaMKII的表达。
四. 讨论
窦房结起搏细胞高度自律性的形成涉及到涉及到多种钠、钾、钙离子通道的协同作用,节律稳定、频率适中的窦性心脏节律则需要各种离子流、信号通路、代谢通路达到一种高度和谐的动态平衡状态。由于心脏电系统具有高度复杂性,一旦这种动态平衡被打破,往往单一靶标的调控是无法使其重新恢复平衡的。动态平衡的重建往往需要对一系列靶点进行“理性的”(rational)的协同调控,以窦性心动过缓的治疗为例,激动功能属性为阳性的靶标、抑制阴性的靶标,或者上调功能属性为阳性靶标的表达、抑制阴性靶标的表达量,简言之即“扶阳抑阴”,则可以起到提升窦房结自律性,加快窦性心率的作用。而对于窦性心动过速的治疗则与上述机制相反,即“扶阴抑阳”。本文通过中医阴阳理论对窦房结起搏机制进行解析和重述,希望能够为中医药治疗窦性心律失常提供更多的思路。
参考文献
[1] 张树生. 对中医阴阳学说的思考与认知[J]. 中医杂志, 2014,55(18):1616-1618.
[2] 李景东,苏冠华,张家明. 对窦房结起搏功能的新认识[J]. 临床心血管病杂志, 2014,03):185-187.
[3] 张晓华,于德洵,钱锋. 病态窦房结综合征中西医研究进展[J]. 中医临床研究,2014,6(15):10-12.
[4] Jones SA,Boyett MR,Lancaster MK. Declining into failure the age-dependent loss of the l-type calcium channel within the sinoatrial node[J]. Circulation,2007,115(10):1183-1190.
[5] 谢晓婷,李泱,张建成. 病窦综合征的离子机制研究进展[J]. 中国心脏起搏与心电生理杂志,2016,(05):1-3.
[6] Noble D,Tsien R. The kinetics and rectifier properties of the slow potassium current in cardiac purkinje fibres[J]. The Journal of physiology,1968,195(1):185-214.
[7] Hauswirth O,Noble D,Tsien R. Adrenaline: Mechanism of action on the pacemaker potential in cardiac purkinje fibers[J]. Science,1968,162(3856):916-917.
[8] DiFrancesco D,McNaughton P. The effects of calcium on outward membrane currents in the cardiac purkinje fibre[J]. The Journal of physiology,1979,289(1):347-373.
[9] Cohen I,Daut J,Noble D. The effects of potassium and temperature on the pace‐maker current,ik2,in purkinje fibres[J]. The Journal of physiology,1976,260(1):55-74.
[10] DiFrancesco D,Ojeda C. Properties of the current if in the sino‐atrial node of the rabbit compared with those of the current ik,in purkinje fibres[J]. The Journal of Physiology,1980,308(1):353-367.
[11] DiFrancesco D. A new interpretation of the pace-maker current in calf purkinje fibres[J]. The Journal of Physiology,1981,314(:359.
[12] DiFrancesco D. A study of the ionic nature of the pace‐maker current in calf purkinje fibres[J]. The Journal of Physiology,1981,314(1):377-393.
[13] DiFrancesco D. The cardiac hyperpolarizing-activated current,i f. Origins and developments[J]. Progress in biophysics and molecular biology,1985,46(3):163-183.
[14] DiFrancesco D. Pacemaker mechanisms in cardiac tissue[J]. Annual review of physiology,1993,55(1):455-472.
[15] Baruscotti M,Bucchi A,DiFrancesco D. Physiology and pharmacology of the cardiac pacemaker (“funny”) current[J]. Pharmacology & therapeutics,2005,107(1):59-79.
[16] Barbuti A,DiFrancesco D. Control of cardiac rate by “funny” channels in health and disease[J]. Annals of the New York Academy of Sciences,2008,1123(1):213-223.
[17] 刘金凤,彭杰,汪艳丽,et al. 窦房结细胞起搏与超极化激活的环核苷酸门控通道关系的研究进展[J]. 中西医结合心脑血管病杂志,2015,14):1622-1625.
[18] 杨一晨,沈法荣,何浪. Hcn4与心脏生物起搏研究进展[J]. 心电与循环,2015,(01):60-62.
[19] Shi W,Wymore R,Yu H,et al. Distribution and prevalence of hyperpolarization-activated cation channel (hcn) mrna expression in cardiac tissues[J]. Circulation research,1999,85(1):e1-e6.
[20] Liu J,Dobrzynski H,Yanni J,et al. Organisation of the mouse sinoatrial node: Structure and expression of hcn channels[J]. Cardiovascular research,2007,73(4):729-738.
[21] Qu J,Plotnikov AN,Danilo P,et al. Expression and function of a biological pacemaker in canine heart[J]. Circulation,2003,107(8):1106-1109.
[22] Guo J,Ono K,Noma A. A sustained inward current activated at the diastolic potential range in rabbit sino-atrial node cells[J]. The Journal of Physiology,1995,483(Pt 1):1.
[23] Rigg L,Terrar DA. Possible role of calcium release from the sarcoplasmic reticulum in pacemaking in guinea‐pig sino‐atrial node[J]. Experimental Physiology,1996,81(5):877-880.
[24] Hüser J,Blatter LA,Lipsius SL. Intracellular ca2+ release contributes to automaticity in cat atrial pacemaker cells[J]. The Journal of Physiology,2000,524(2):415-422.
[25] Ju YK,Allen DG. Intracellular calcium and na+‐ca2+ exchange current in isolated toad pacemaker cells[J]. The Journal of physiology,1998,508(1):153-166.
[26] Maltsev VA,Lakatta EG. Synergism of coupled subsarcolemmal ca2+ clocks and sarcolemmal voltage clocks confers robust and flexible pacemaker function in a novel pacemaker cell model[J]. American Journal of Physiology-Heart and Circulatory Physiology,2009,296(3):H594-H615.
[27] Lakatta EG,Maltsev VA,Vinogradova TM. A coupled system of intracellular ca2+ clocks and surface membrane voltage clocks controls the timekeeping mechanism of the heart’s pacemaker[J]. Circulation research,2010,106(4):659-673.
[28] Accili EA,Proenza C,Baruscotti M,et al. From funny current to hcn channels: 20 years of excitation[J]. News in Physiological Sciences,2002,17(1):32-37.
[29] Altomare C,Baruscotti M,Bucchi A,et al. Allosteric voltage-dependent gating of hcn channels[J]. Biophysical Journal,2001,80(1):241A-241A.
[30] Altomare C,Terragni B,Brioschi C,et al. Heteromeric hcn1-hcn4 channels: A comparison with native pacemaker channels from the rabbit sinoatrial node[J]. Journal of Physiology-London,2003,549(2):347-359.
[31] Randall A,Tsien R. Contrasting biophysical and pharmacological properties of t-type and r-type calcium channels[J]. Neuropharmacology,1997,36(7):879-893.
[32] Lee J-H,Daud AN,Cribbs LL,et al. Cloning and expression of a novel member of the low voltage-activated t-type calcium channel family[J]. The Journal of neuroscience,1999,19(6):1912-1921.
[33] Mangoni ME,Traboulsie A,Leoni A-L,et al. Bradycardia and slowing of the atrioventricular conduction in mice lacking cav3. 1/α1g t-type calcium channels[J]. Circulation research,2006,98(11):1422-1430.
[34] Maltsev VA,Lakatta EG. Cardiac pacemaker cell failure with preserved if,ical,and ikr: A lesson about pacemaker function learned from ischemia-induced bradycardia[J]. Journal of molecular and cellular cardiology,2007,42(2):289-289.
[35] Janvier N,Boyett M. The role of na-ca exchange current in the cardiac action potential[J]. Cardiovascular research,1996,32(1):69-84.
[36] Wongcharoen W,Chen Y-C,Chen Y-J,et al. Effects of a na+/ca2+ exchanger inhibitor on pulmonary vein electrical activity and ouabain-induced arrhythmogenicity[J]. Cardiovascular research,2006,70(3):497-508.
[37] Lei M,Brown HF. Two components of the delayed rectifier potassium current,ik,in rabbit sino‐atrial node cells[J]. Experimental Physiology,1996,81(5):725-741.
[38] 刘丽,李泱,卢才义. Kcne对心肌细胞钾离子通道的调控及与遗传性心律失常[J]. 中国心脏起搏与心电生理杂志,2016,03):260-263.
[39] Lei M,Honjo H,Kodama I,et al. Characterisation of the transient outward k+ current in rabbit sinoatrial node cells[J]. Cardiovascular research,2000,46(3):433-441.
[40] Bursac N,Papadaki M,White JA,et al. Cultivation in rotating bioreactors promotes maintenance of cardiac myocyte electrophysiology and molecular properties[J]. Tissue engineering,2003,9(6):1243-1253.
[41] Takano M,Noma A. Distribution of the isoprenaline-induced chloride current in rabbit heart[J]. Pflügers Archiv,1992,420(2):223-226.
[42] Miake J,Marbán E,Nuss HB. Functional role of inward rectifier current in heart probed by kir2. 1 overexpression and dominant-negative suppression[J]. Journal of Clinical Investigation,2003,111(10):1529-1529.
[43] Grover GJ. Pharmacology of atp-sensitive potassium channel (katp) openers in models of myocardial ischemia and reperfusion[J]. Canadian journal of physiology and pharmacology,1997,75(4):309-315.
[44] Bettahi I,Marker CL,Roman MI,et al. Contribution of the kir3. 1 subunit to the muscarinic-gated atrial potassium channel ikach[J]. Journal of Biological Chemistry,2002,277(50):48282-48288.
[45] Musa H,Lei M,Honjo H,et al. Heterogeneous expression of ca2+ handling proteins in rabbit sinoatrial node[J]. Journal of Histochemistry & Cytochemistry,2002,50(3):311-324.
[46] Smits JP,Koopmann TT,Wilders R,et al. A mutation in the human cardiac sodium channel (e161k) contributes to sick sinus syndrome,conduction disease and brugada syndrome in two families[J]. Journal of molecular and cellular cardiology,2005,38(6):969-981.
[47] Coppen SR,Kodama I,Boyett MR,et al. Connexin45,a major connexin of the rabbit sinoatrial node,is co-expressed with connexin43 in a restricted zone at the nodal-crista terminalis border[J]. Journal of Histochemistry & Cytochemistry,1999,47(7):907-918.
[48] 侯平,刘宁,张明雪,et al. 参仙升脉口服液对缓慢性心律失常大鼠 camp/cgmp,na^+-k^+-atp 酶的影响[J]. 辽宁中医杂志,2012,39(2):360-361.
[49] Jing H,Luo L,Li H,et al. Ephedrine controls heart rhythms by activating cardiac iks currents[J]. Journal of cardiovascular pharmacology,2010,55(2):145-152.
[50] 张水馨.基于 ca~(2+)-cam-camkⅱ 信号传导通路的麻黄附子细辛汤对 cam/camkⅱ 的影响. 北京中医药大学,2014.
[51] 刘艳,章诗伟,周兰,et al. 乌头类生物碱对心肌的毒性作用及分子毒理学研究进展[J]. 中国法医学杂志,2009,06):398-401.
[52] 黄悦. 稳心颗粒基础及临床研究进展[J]. 首都医药,2014,10(06):15-17.
[53] 李玉成. 步长稳心颗粒治疗窦性心动过速的疗效观[J]. 中国实用医药,2010,(08):176.
[54] 刘玉红,舒桂兰. 稳心颗粒治疗更年期室上性心动过速21例[J]. 中西医结合心脑血管病杂志,2006,(11):1011.
[55] 吕晓东,张秀平. 步长稳心颗粒治疗肺心病引起窦性心动过速的临床观察[J]. 中国实用医药,2013,(33):147.
[56] 王锦乾,阮爱兵,吕爱玲. 步长稳心颗粒治疗肺心病伴室上性心动过速50例[J]. 中医杂志,2005,(11):842.
[57] 徐利贵,周伟平,王金亮,et al. 步长稳心颗粒治疗甲亢性心动过速的疗效观察[J]. 中国实用医药,2007,(08):54-55.
[58] 徐云. 步长稳心颗粒治疗肺心病引起窦性心动过速的疗效观察[J]. 中西医结合心血管病电子杂志,2015,(27):75-76.
[59] Chen Y,Li Y,Guo L,et al. Effects of wenxin keli on the action potential and l-type calcium current in rats with transverse aortic constriction-induced heart failure[J]. Evidence-based complementary and alternative medicine : eCAM,2013,2013.
[60] Burashnikov A,Petroski A,Hu D,et al. Atrial-selective inhibition of sodium-channel current by wenxin keli is effective in suppressing atrial fibrillation[J]. Heart Rhythm,2012,9(1):125-131.
[61] Wang X,Wang X,Gu Y,et al. Wenxin keli attenuates ischemia-induced ventricular arrhythmias in rats: Involvement of l-type calcium and transient outward potassium currents[J]. MOLECULAR MEDICINE REPORTS,2013,7(2):519-524.
[62] Xing Y,Gao Y,Chen J,et al. Wenxin-keli regulates the calcium/calmodulin-dependent protein kinase ii signal transduction pathway and inhibits cardiac arrhythmia in rats with myocardial infarction[J]. Evidence-based complementary and alternative medicine:eCAM,2013,2013.
基金项目:科技部国家国际合作专项(2013DFA31620) “抗心律失常复方中药稳心颗粒的药效物质基础和分子作用机理的合作研究”。